L'organisation et la diversité du vivant
PLAN DU COURS
I - Définir la biodiversité actuelle et passée
II - Réaliser des groupes au sein des être-vivants et établir des parentés
III - Créer et faire évoluer la biodiversité : dérive génétique et sélection naturelle
Les scientifiques ont démontré que le monde vivant a subi de grandes transformations tout au long de son histoire et les espèces actuelles ne représentent qu'1% des espèces ayant vécu sur Terre. L'étude des fossiles en apporte la preuve.
Comment mettre en évidence la modification du monde vivant au cours des temps géologiques ?
I - Définir la biodiversité actuelle et passée
Pour prouver que le monde vivant a fortement évolué au cours des temps nous allons étudier 2 écosystèmes:
- une image reconstituant la forêt houillère du Carbonifère (300Ma; Ma = millions d'années)
- une photo de la forêt tropicale guyanaise actuelle.
Définition d'écosystème : un écosystème est l'ensemble formé par une association d'êtres vivants (= biocénose) interagissant avec leur environnement biologique, géologique,édaphique, hydrologique, climatique, etc. (= le biotope). Pour résumer: l'écosystème = biotope + biocénose.



Établissons un tableau comparant ces 2 écosystèmes:
|
Écosystèmes |
Végétaux | Animaux | Milieux de vie | |||||
| Plantes à fleurs | Plantes sans fleurs | Arthropodes | Oiseaux | Reptiles | Mammifères | Amphibiens | Chaud et humide | |
| Actuel (GUYANE) | Forte | Faible | Forte | Moyenne | Faible | Faible | Faible | OUI |
| Passé (FORET HOUILLERE) | Absentes | Forte (fougères, prêles) | Forte (blattes, libellules) | Absents | Absents | Absents | Faible | OUI |
On peut conclure qu'il y a 300Ma la biodiversité était essentiellement constituée de prêles et fougères, de cycadales et de conifères. Les plantes à fleurs n'étaient pas encore apparues.
Elles apparaissent au Crétacé (130Ma) et depuis la biodiversité végétale a fortement augmenté (voir forêt guyanaise).
La vie serait apparue il y a plus de 3,8Ga mais c'est depuis 600Ma que de nombreux fossiles sont présents en grande quantité dans les roches sédimentaires du même âge. En étudiant les différents
fossiles, leur nombre et le nombre d'espèces que contiennent les roches sédimentaires on peut reconstituer l'évolution du nombre de familles d'animaux depuis 600Ma. Les résultats sont disponibles
dans le graphique ci-dessous :

Grâce à cette étude on découvre que 5 grandes crises biologiques ont touché la Terre amenant à chaque fois la disparition d'une grande quantité de familles d'animaux.
Les principales causes à l’origine des extinctions de masse sont:
- les cataclysmes géologiques (éruptions volcaniques, variations du niveau marin, glaciation),
- les chutes d’astéroïdes,
- des changements climatiques,
- la prédation (cas du dodo de l’île Maurice),
- la compétition (certaines espèces introduites se multiplient rapidement,
au détriment d’espèces locales),
- des maladies, virus, des causes génétiques,
- la disparition ou l’extinction d’une ressource dont dépendait l’espèce pour sa survie (ex: panda et bambous) ou sa reproduction : pollinisateur, proie…
BILAN: au cours des temps géologiques les espèces peuplant les différents écosystèmes ont changé: des espèces sont apparues, d'autres ont disparu.
Les espèces actuelles ne représentent qu'une infime proportion des espèces ayant vécu sur Terre. Les variations de l'environnement (climat, catastrophes naturelles...) sont les principales causes des modifications de la biodiversité.
La biodiversité peut s'appréhender à différentes échelles :
- la diversité des écosystèmes à l'échelle mondiale,
- la diversité des espèces,
- la diversité génétique (allèles).
- la diversité des écosystèmes à l'échelle mondiale. Un biome est constitué par des grands types de formations végétales caractéristiques d'une grande zone climatique de la planète. A ces associations végétales (=phytocénoses) s'ajoutent des associations animales (= biocénoses) dont le tout forme un biome.
Exemple: la savane africaine dans laquelle on retrouve les lions, girafes... et les grandes herbacées et les acacias...
Attention un biome ne constitue pas un écosystème, c'est une zone où les conditions du milieu permettent aux espèces des interactions entre elles et ce milieu.

- la diversité des espèces: sur Terre on retrouve un très grand nombre d'espèces (environ 1,8 millions recensées à l'heure actuelle) mais les estimations sont bien supérieures: il y aurait entre 2 et 10 millions d'espèces sur Terre.

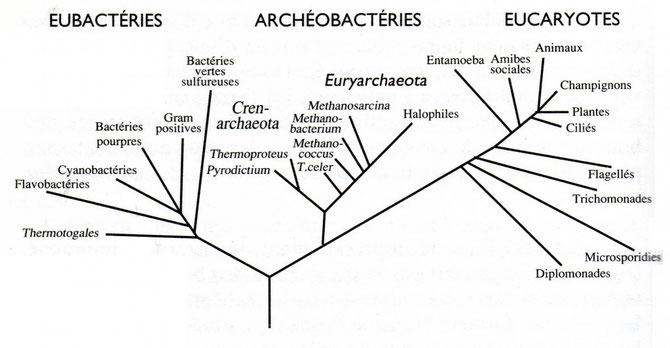
L'arbre phylogénétique ci-dessous détaille les 3 grands embranchements de la vie. Les différentes espèces qui les composent sont issues d'un ancêtre commun appelé LUCA (Last Universal Common Ancestor). Cet ancêtre aurait possédé une membrane et un cytoplasme entourant une molécule d'ADN. Car ce sont les seuls points communs que l'on retrouve chez tous les êtres-vivants actuellement !
A cela s'ajoute que la Vie est constituée par 6 principaux éléments atomiques: C,H,N,O,P,S.
Une étude sur LUCA : lien.
Ernst Mayr a donné une définition d'une espèce :
Définition: une espèce correspond à un groupe de populations naturelles semblables dans lequel les individus peuvent échanger du matériel génétique et avoir une descendance féconde (par reproduction la plupart du temps). Une espèce est séparée d'une autre espèce par des mécanismes d'isolement reproducteurs: c'est-à-dire qu'une espèce ne peut pas se reproduire avec une autre espèce et donner des descendants féconds (voir exemples classiques des Tigrons, Ligres...).
Parmi toutes les espèces vivantes, on défini comme zones critiques ou hot spots ou points chauds les zones qui possèdent une très grande richesse de biodiversité menacée par l'activité humaine. Ces zones représentent 15% de la surface terrestre mais abritent un nombre très important d'espèces.
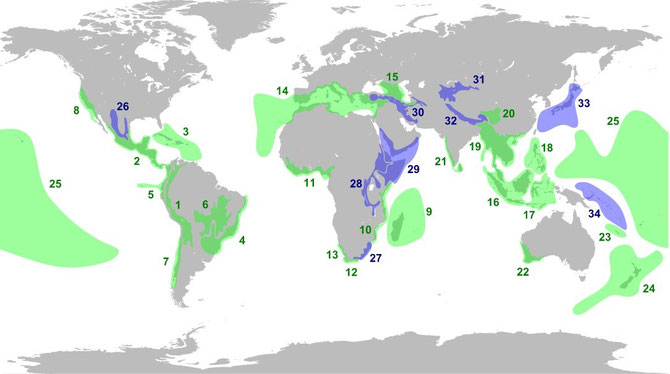
Amérique
Province floristique de Californie (8 - Bande côtière entre le l'extrême nord-ouest du Mexique et le Nord de la Californie)
Bois de pins et chênes de Madrean (26 - Sud-ouest des États-Unis et Nord du Mexique)
Amérique centrale (2 - Du sud du Mexique au Panama)
Îles des Caraïbes (3)
Tumbes-Chocó-Magdalena (5 - Bande côtière entre le Panama et Nord du Pérou)
Andes tropicales (1 - Du Venezuela au nord-ouest de l'Argentine)
Cerrado (6 - Est du Brésil, Bolivie et Paraguay)
Forêt atlantique (4 - Sud-est du Brésil, Nord du Paraguay, extrême nord-est de l'Argentine)
Forêts pluviales tempérées valdiviennes (7 - Centre et Sud du Chili)
Europe
Bassin méditerranéen (14 - Bassin méditerranéen et Macaronésie: Canaries, Acores, Madère, Cap-Vert)
Caucase (15 - Géorgie, Arménie, Azerbaïdjan, Russie, Turquie et Iran)
Afrique
Corne africaine (29 - Éthiopie, Somalie, Kenya, Érythrée, Djibouti, Yémen et Oman)
Afromontane orientale (28 - Sud-ouest de la Péninsule Arabique et montagnes entre l'Éthiopie au Zimbabwe)
Forêt guinéenne de l'Ouest africain (11 - De la Guinée au Cameroun)
Forêts côtières d'Afrique orientale (10 - Bande côtière entre la Somalie et le Mozambique)
Madagascar et îles de l'Océan Indien (9 - Madagascar, Mascareignes, Comores, Seychelles)
Karoo succulent (13 - Ouest de l'Afrique du Sud et Sud-ouest de la Namibie)
Maputaland-Pondoland-Albany (27 - Côte est de l'Afrique du Sud)
Région floristique du Cap (12 - Sud-ouest de l'Afrique du Sud)
Asie
Désert irano-anatolien (30 - Arménie, Azerbaïdjan, Turquie, Géorgie, Turkménistan, Iran et Irak)
Montagnes d'Asie centrale (31 - Kazakhstan, Kirghizstan, Tadjikistan, Ouzbékistan, Chine, Afghanistan et Turkménistan)
Himalaya (32 - Chine, Inde, Bhoutan, Pakistan, Afghanistan)
Montagnes de la Chine Occidentale (20)
Japon (33)
Ghâts occidentaux (21 - Ouest de l'Inde et Sri Lanka)
Indo-Burma (19 - Du Bangladesh à l'extrême sud-est de la Chine et du nord de la Birmanie au Sud de la Thaïlande)
Sundaland (16 - Péninsule Malaise, Singapour, Brunei, Bornéo, Sumatra, Java et Bali)
Philippines (18)
Wallacea (17 - Sulawesi, Petites îles de la Sonde, Moluques et Timor)
Océanie
Mélanésie orientale (34 - Archipel Bismarck, îles Salomon et Vanuatu)
Polynésie et Micronésie (25 - 4500 îles allant des Palaos à l'île de Pâques et de Hawaï jusqu'aux Fidji)
Nouvelle-Calédonie (23)
Sud-Ouest de l'Australie (22)
Nouvelle-Zélande (24)
- la diversité génétique au sein d'une espèce (= diversité allélique):

Un texte écrit par Hervé Le Guyader, grand chercheur systématicien :
La biodiversité désigne toutes les variations du monde vivant. Cette diversité biologique est évidente lorsqu’on imagine la quantité fantastique d’espèces différentes (découvertes ou encore inconnues) qui vivent ou ont vécu sur Terre. C’est la biodiversité spécifique. Au sein de chaque espèce, on constate un autre niveau de biodiversité : la biodiversité génétique, c’est à dire la diversité des allèles d’un individu à l’autre. Ainsi, différents humains posséderont les mêmes gènes, mais pas les mêmes allèles de ces gènes. Le troisième niveau est la biodiversité des écosystèmes. Il correspond à la diversité des milieux de vie, incluant les espèces qui les habitent et toutes les relations qu’elles ont entre elles (qui mange qui, qui parasite qui, etc…).
Hervé Le Guyader, chercheur en systématique et évolution

Les scientifiques estiment à environ 10 millions le nombre d'espèces vivant sur Terre (pour environ 2 millions d'espèces découvertes et classées). Ces êtres-vivants sont très variés de par leur morphologie ou leur milieu de vie mais aussi au niveau de leurs fonctions alimentaire, respiratoire, reproductrice... Pourtant ces individus très différents proviennent tous d'un ancêtre commun (le fameux LUCA) et peuvent être classés dans des groupes bien définis.
II - Réaliser des groupes au sein des être-vivants et établir des parentés
La biodiversité regroupe l'ensemble des espèces vivantes en considérant la diversité des écosystèmes qu'elles habitent, leurs différences spécifiques (visibles entre espèces) et leurs diversité infraspécifique (variable suivant les différences entre leurs allèles).
Depuis le collège vous savez:
- classer en groupes emboîtés les taxons* qui partagent des caractères communs
- les espèces comme les groupes (ou taxons) apparaissent, disparaissent au cours des temps géologiques.
- une espèce nouvelle présente des caractères ancestraux et aussi des caractères nouveaux par rapport à une
espèce antérieure dont elle serait issue.
- utiliser le logiciel Phylogène (normalement...).
*Taxon: ensemble d'organismes vivants ou disparus possédant des caractère taxinomiques (de classification) identiques. Ex: le taxon des mammifères possédant des poils et des mamelles permettant d'allaiter les jeunes.
Au sein des Vertébrés (42 000 espèces), on trouve des animaux bien différents (grenouille, souris, serpents...) au niveau de leur plan d'organisation* ! De tout temps l'Homme à chercher à classer les espèces et établir des liens entre elles.
* plan d'organisation : disposition de ses différents éléments constitutifs (axes de polarité et place des différents organes).
Étudions la morphologie de la grenouille et de la souris, 2 vertébrés très différents :


L'étude morphologique de ces 2 animaux montre des ressemblances:
- des axes de polarité identiques (antéro-postérieur = d'avant en arrière; dorso-ventral = du dos vers le ventre; et gauche-droite);
- au niveau du nombre de membres (4);
- présence d'un tête, d'une bouche et de 2 yeux en position antérieure.
Voyons si au niveau anatomique des ressemblances existent aussi:
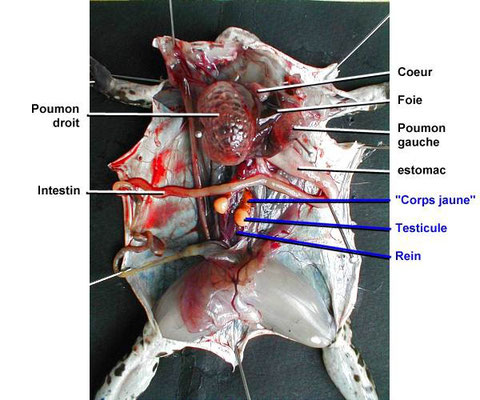

Sur le schéma ci-dessous nous avons résumé les principaux organes constituant l'appareil respiratoire et l'appareil digestif de ces animaux.

Classer les espèces sur un arbre phylogénétique :
pour établir des parentés entre des espèces on compare des caractères homologues* (morphologiques, anatomiques, moléculaires…). On établit une matrice des caractères (tableau à double entrée résumant les observations réalisées chez les groupes étudiés), et on construit un arbre
phylogénétique qui traduit les relations de parenté. Chaque nouvelle branche doit être justifiée par l’apparition d’une nouveauté évolutive (ex: apparition des mamelles chez les mammifères). Un nœud correspond à la population des derniers ancêtres communs à partir de laquelle ont émergé 2 groupes différant par leurs caractéristiques.
* caractère homologue: caractère hérité d'un ancêtre commun.

Autre méthode de classification: les groupes emboités :
les organismes vivants sont donc liés entre eux par des ancêtres communs. On dit alors qu'il existe des liens de parenté entre les êtres vivants. On peut ainsi classer les espèces par groupes (ou taxons) en fonction des caractères qu'elles présentent. En comparant les groupes entre eux on peut alors définir des caractères qu'ils partagent ou qui les séparent. On constitue ainsi des matrices de caractères traduisant des liens de parenté entre les espèces et groupes.
On peut ensuite représenter ces liens de parenté sous la forme de groupes emboîtés, reliant ainsi les taxons entre eux :



BILAN:
Il existe des parentés entre les espèces vivantes qui composent la biodiversité. Ces parentés définissent des groupes d’êtres vivants. Le groupe des Vertébrés est défini par la possession de caractères exclusifs : la colonne vertébrale, les deux axes de polarité et la symétrie bilatérale. Les parentés d’organisation des espèces d’un groupe suggèrent qu’elles descendent toutes d’un ancêtre commun.
III - Créer et faire évoluer la biodiversité : dérive génétique et sélection naturelle
Les caractères définissant une espèce sont déterminés par les gènes. Chaque espèce possède un nombre précis de gène et un nombre fixe de chromosomes. Chez l'Homme, il existe 46 chromosomes répartis en 23 paires de taille identique (voir cours de 3ème) et portant les mêmes gènes (environ 20 000). Mais ces gènes peuvent posséder des variations appelées les allèles. On appelle polymorphisme génétique toutes les différences entre 2 individus que l'on peut observer au niveau de leur ADN et des produits de leurs gènes.
Le système de groupe sanguin ABO a été découvert au début du XXe siècle par Landsteiner. L’étude de ce système, pour des besoins transfusionnels, démontra très tôt l’existence des variations génétiques parmi les populations humaines.
Par exemple pour le caractère "groupe sanguin" il existe 1 gène sur le chromosome n°9 et 3 allèles de ce gène (c'est-à-dire 3 versions différentes): les allèles A, B et O permettent de créer les 4 types de groupes sanguins (voir tableau ci-dessous).

La distribution des allèles du système dans le monde a été largement étudiée.
L'étude des fréquences de ces allèles montre qu'ils ne sont pas homogènes dans les populations. Au contraire, ils sont présents à certains endroits précis dans le monde (voir cartes ci-dessous).




L'étude de ces 4 cartes montre que la répartition des différents allèles des groupes sanguins n'est pas homogène au sein de la population humaine. Par exemple l'allèle B correspondant au groupe sanguin B est sur-représenté en Inde : cela pourrait s'expliquer par les faible migrations au sein de ce pays, le faible apport d'allèles provenant de personne migrant dans ce pays et un choix local quant à son partenaire de reproduction (les indiens ont des enfants entre eux et au sein des mêmes castes).
Autre exemple: la répartition des groupes ABO chez les Indiens d’Amérique, révèle une fréquence extrêmement élevée de l’allèle O. Pour expliquer l’absence des groupes A et B sur le continent Américain, il a été proposé un effet fondateur lié au fait qu’un petit groupe d’Homo Sapiens de groupe O aurait franchi le détroit de Béring il y a 15 000 ans et se seraient installé en Amérique du Nord.
La dérive génétique:
La dérive génétique correspond à l'évolution de la fréquence des allèles au sein d'une population. La fréquence des allèles d'une population est impossible à prévoir car la dérive génétique correspond à l'évolution aléatoire des fréquences alléliques (= des allèles) au sein d'une population.
La répartition des allèles lors de la formation des gamètes ou de la fécondation se fait au hasard (cours de 3ème). Ainsi seule une partie du patrimoine génétique est transmise à la descendance. Cela engendre des variations aléatoires de la fréquence des allèles d'un même gène dans la population. Tout allèle peut donc disparaître ou se répandre d'autant plus vite que la population d'origine est faible. Cette variation est appelée dérive génétique. Elle est souvent associée à un effet fondateur (voir exemple des Huttérites ci-dessous)

L'effet fondateur (à limite du programme de 2nde) :
Pour comprendre la répartition de certains allèles dans une population étudions une population humaine : les Huttérites. Ce sont les membres d'une secte qui, persécutés en Europe, se sont installés en Amérique du Nord au dix-neuvième siècle. Comme les Amish, ils ont établi une série de colonies d'une centaine d'individus dans lesquelles ils vivent en autarcie sans se marier avec les personnes étrangères.
Les Huttérites présentent des fréquences alléliques ABO qui sont très différentes des moyennes constatées dans les populations européennes et nord-américaines (ex: 35% de A chez les Huttérites contre 2% pour les Nord-Américain). Cela provient du fait que chaque colonie a été fondée à partir d'un petit nombre d'individus dont les fréquences des allèles étaient très différentes des fréquences moyennes des populations européennes dont ils sont issus. Cela induit une dérive génétique très forte et très rapide !
Considérons que 2 populations (les Huttérites d'Europe quittent les Européens) se séparent à un moment donné. Elles auront initialement les mêmes fréquences alléliques (1ère population du schéma), mais elles vont connaître des divergences des fréquences alléliques (se modifiant aléatoirement) au cours du temps.
La variabilité génétique de l'espèce va augmenter, ceci d'autant plus qu'elle sera composée de petites populations isolées les unes des autres. La fréquence allélique d'un groupe migrant n'est souvent pas représentative de la population dont il est issu. C'est l'effet fondateur.

BILAN:
la diversité des allèles d’un gène est un des aspects de la biodiversité. Elle se manifeste au sein des populations qui peuvent présenter des diversités alléliques très différentes.
La dérive génétique est l’évolution génétique d’une population causée par des phénomènes aléatoires, impossibles à prévoir (d’où le mot dérive !). Elle est une conséquence de la reproduction sexuée des individus. La dérive génétique conduit, pour la population, à une perte de diversité génétique. Elle se produit de façon plus marquée lorsque l’effectif de la population est faible (= son effet est plus fort).
La sélection naturelle :
la dérive génétique n'est pas la seule force évolutive modifiant la fréquence des allèles d'une population. La sélection naturelle joue aussi un rôle très importante.
La Phalène du Bouleau, Biston betularia, est un papillon de nuit, commun en Europe dont la coloration varie du gris clair au noir. Il y a 150 ans, les populations de l’espèce étaient presque exclusivement constituées de la forme « typica », aux ailes gris clair légèrement mouchetées de noir.
En 1848, un individu d’une forme mélanique « carbonaria », uniformément noir, fut collecté près de Manchester. En 1950, la forme mélanique représentait 90% de la population dans cette région.
Ce phénomène (apparition d’individus très sombres dans la deuxième moitié du 19eme siècle et augmentation rapide de leur fréquence) fut observé chez de nombreuses autres espèces de papillons de nuit, ainsi que chez différentes espèces d’insectes, d’araignées, et même d’oiseaux dans les environs d’autres villes industrielles telles que Birmingham et Liverpool.


Au milieu du XIXème siècle, la forme typica était largement majoritaire dans les populations anglaises (carte de gauche). En 1848 on a capturé le premier individu carbonaria dans la région de Manchester. La fréquence de cette forme s'est alors accrue rapidement et les individus mélaniques ont ensuite été observés dans les autres régions industrielles de l'Angleterre (carte de droite).

Dès le XIXème siècle, on corréla l’augmentation de la fréquence de la forme mélanique à la pollution industrielle. Celle-ci contribue à la disparition des lichens sur les bouleaux et au noircissement de ces arbres.
On suggéra que les individus « typica », posés sur des bouleaux clairs couverts de lichens, étaient mieux dissimulés vis-à-vis des oiseaux prédateurs que les individus « carbonaria ». Sur des bouleaux noircis par la suie et dépourvus de lichens, c'est l'inverse qui se produisait. Les deux formes seraient alors consommées de façon différente selon le type de support, la forme mélanique étant avantagée sur des arbres noircis.
Ainsi selon l'environnement, l'une des formes de papillons, mieux
« camouflée », échappe à ses prédateurs. Ce papillon aura plus de chance de se reproduire et il transmettra sa couleur à ses descendants: ainsi la fréquence des allèles qu'il possède augmente dans la population et l'une des formes devient rapidement majoritaire. C'est la sélection naturelle : une mutation de la couleur du papillon, phénomène aléatoire, confère aux individus qui en sont porteurs un avantage sélectif; ceux-ci se répandent dans la population. Inversement les mutations défavorables ont tendance à être éliminées, car les individus qui en héritent survivent plus difficilement et se reproduisent moins.

Du fait de certaines conditions de milieu, certains individus laissent plus de descendants que d’autres. Ce mécanisme de « tri » des individus par l’environnement se nomme sélection naturelle. Il est totalement dépendant des conditions environnementales.
Dérive génétique et sélection naturelle en modifiant la fréquence des allèles au sein des populations peuvent conduire à l’apparition de nouvelles espèces (s’il n’y a pas d’échanges d’allèles par migration).
Lien pour réviser la structure et les organites d'une cellule : LIEN.

Sites:
- Les écologistes de L'Euzière: LIEN.
- Enseigner la classification des vertébrés: LIEN
- Une page sur LUCA (Last Universal Common Ancestor): LIEN.
Pour plus d'informations sur la reconnaissance des plantes de la garrigue suivre les liens vers les livres numériques (et très bien faits) de Marcel Vaillaud (très belles photos) sur la garrigue:
- les arbres: LIEN,
- les arbustes: LIEN,
- les lianes: LIEN,
- bulbes et rhizomes: LIEN,
- les adaptations à la sécheresse: LIEN,
- les lichens: LIEN.
Écrire commentaire